|
|
|
|
|
|
This article may be reprinted free of charge provided 1) that there is clear attribution to the Orthomolecular Medicine News Service, and 2) that both the OMNS free subscription link http://orthomolecular.org/subscribe.html and also the OMNS archive link http://orthomolecular.org/resources/omns/index.shtml are included. FOR IMMEDIATE RELEASE
Vitamin C Levels in Critically Ill Covid-19 Patients
by Michael Passwater(OMNS July 18, 2021) From the COVID-19 pandemic, we continue to learn about the critical importance of maintaining adequate levels of essential nutrients. When the body is under stress from an illness such as an infection, merely eating an excellent diet may not provide sufficient nutrients to support the immune system. To stave off a fast-moving infection may require higher levels of essential vitamins and minerals. Vitamin C has an essential role in empowering the immune system. Its oxidized form can be recycled by red blood cells (erythrocytes), but a fast-moving illness can overpower this system, causing vitamin C levels to precipitously drop. A recent study in Spain measured plasma vitamin C levels in 67 critically ill hospitalized adult COVID-19 patients meeting the Berlin criteria for Acute Respiratory Distress Syndrome (ARDS). [1] The results fell into 3 categories: undetectable (<0.1 mg/dL), low (0.1 - 0.4 mg/dL), and "normal" (0.4 - 2 mg/dL). 12 (18%) patients had undetectable plasma vitamin C, 43 (64%) patients had low levels of plasma vitamin C (mean for this group was 0.14 mg/dL with a standard deviation of 0.05), and 12 (18%) patients had vitamin C levels within the normal range (mean for this group was 0.59 mg/dL with a standard deviation of 0.18). In summary, 82% of patients had low or undetectable plasma vitamin C levels, and 18% had values within the reference range, mostly on the low side of the reference range. (Riordon Clinic Bio-Center Laboratory has an established reference range of 0.6 - 2.0 mg/dL for plasma vitamin C). A smaller study of 18 adult COVID-19 patients with ARDS found similar results: 17 (94%) patients had undetectable plasma vitamin C, and 1 (6%) patient had a plasma vitamin C level of 0.24 mg/dL. [2] The assay used in this study had a lower limit of detection of 0.15 mg/dL, above the mean of the low level group in the first study. Finding low levels of vitamin C in critically ill patients is not new, and has been reported in a variety of studies over the last several decades. In 2017, a study of 44 critical care patients receiving recommended amounts of enternal and parenternal vitamin C (125 +/- 88 mg/day, max 448 mg/day) showed 70% of patients had vitamin C deficiency. [3] Among septic shock patients, 90% had vitamin C deficiency. Borrelli et al published findings in 1996 showing that the lower the plasma ascorbic acid level in septic patients the greater the risk of organ failure and death. [4] Even in presumed healthy people in the USA, vitamin C deficiency is found. In 2003-2004, NHANES samples from noninstitutionalized civilians found a vitamin C deficiency prevalence of 7.1% +/- 0.9%. [5] This was a 44% reduction in vitamin C deficiency from the 1988 - 1994 national study. Smoking and low income were associated with higher rates of vitamin C deficiency. People in a deficient state can avoid acute illness for a time, but have impaired capacity to respond to infections and other stress challenges. Why do critically ill people require more vitamin C to maintain adequate levels of plasma vitamin C?
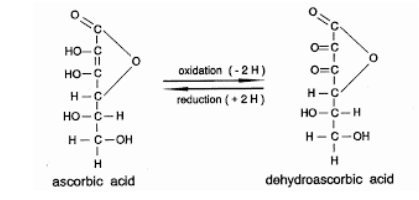
In addition to maintaining antioxidant capacity, RBCs are responsible for the management of the three gases of life, O2, CO2, and NO, throughout the body. [10] RBCs (erythrocytes), are produced from erythroid precursor cells in the bone marrow, and circulate for approximately four months. They are biconcave discs, with very flexible membranes to allow them to flow smoothly throughout the body's 60,000 miles of blood vessels. Capillaries in the body's extremities become so narrow that the RBCs flow single file, underscoring the necessity of cell membrane flexibility. New research reveals that RBC membrane components, interferon, and selenoproteins are targets of the SARS-CoV-2 virus, and along with NAD are all depleted by the virus. [11-15] In addition to GLUT1 receptors, RBC membranes also can express ACE2 receptors, which are well established as a cellular entry point for the SARS-CoV-2 virus. CD147 and the RBC structural protein Band3 have also been shown to serve as attachment points for the virus. Mature RBCs do not have a nucleus and cannot support viral replication. However viral attachment and entry can disrupt the RBC's ability to transport and transfer oxygen to tissues, as well as destroy selenoproteins which in turn disrupts DHAA - AA recycling. RBC membrane disturbances and loss of antioxidant capacity results in a more spherical and less flexible RBC, and oxidation causes phosphatidyl serine and other lipids to flip from the inner side of the membrane to the outer side of the membrane. These changes inhibit the RBC from bending and twisting to travel through the small capillaries of the circulatory system, and accelerate the RBC's clearance from circulation by the reticuloendothelial system monocytes in the spleen and liver. Immature RBC precursor cells have a nucleus, numerous ACE2 receptors, and can support viral replication. Invasion of these cells by the SARS-CoV-2 virus is even more damaging. Release of RBC precursor cells into the blood stream in response to hypoxia, can intensify the disease by causing immunosuppression and serving as a rich source of selenocysteine and other nutrients for the rapidly replicating virus. The virally induced structural, functional, and metabolic damage to RBCs helps explain cases of COVID-19 presenting with hypoxia disproportional to the degree of pneumonia present. In addition to elucidating the interactions of SARS-CoV-2 with the RBCs and RBC precursor cells, recent genetics, proteomics, metabolomics, and lipidomics research has identified specific interactions leading to interferon and selenoprotein destruction and suppression. These studies have also identified nicotinamide phosphoribosyltransferase, nicotinamide, and nicotinamide riboside as therapeutic options to boost innate immunity and counteract NAD depletion by the virus. Importance of adequate niacin, glutathione/cysteine (NAC and alpha lipoic acid), and selenium The findings of recent studies on the effect of nutrient deficiencies in COVID-19 add empirical evidence in support of hypotheses published early in the pandemic. In March of 2020, Yufang Shi and team in China recommended the use of niacin (vitamin B3) whenever lung damage was detected by CT scan. [16] Miller, Wentzel, and Richards in South Africa pointed to the importance of NAD+ deficiency. [17] Over a decade ago, Ethan Will Taylor proposed the oxidative stress-induced niacin sink (OSINS) model for HIV, another RNA virus. [18] Taylor, along with Hiffler, Vavougios, Polonikov and others also suggested glutathione and selenium as central in the etiology of SARS-CoV-2 disease. [19-21] Additionally, a German study showed an inverse association between COVID-19 mortality or severe illness and selenium and selenoprotein P levels. [22] And in the USA, two cases of severe COVID-19 were successfully treated with oral and intravenous glutathione, N-acetyl-cysteine (NAC), and lipoic acid have been published. [23] ConclusionSARS-CoV-2 is an RNA virus capable of causing systemic, life-threatening disease in humans. Severe disease is characterized by hyper-inflammation, hyper-coagulation, and hypoxia. The virus produces proteins that knockdown two major pillars of the innate immune system, interferon and selenoproteins. Selenoprotein knockdown impairs antioxidant capacity and hemostasis (anticoagulation and clotting). The virus also damages RBC structure, which combined with loss of antioxidant capacity, impairs management of oxygen, carbon dioxide, and nitric oxide throughout the body. Additionally, consumption of NAD by the virus depletes cells of a vital energy source. Restoring and maintaining healthy levels of ascorbic acid, selenocysteine, vitamin D, and NAD is critical in the battle against SARS-CoV-2. In treatment, as well as in research, it is important to remember that nutrients do not work optimally alone. Selenium, vitamin D, magnesium, and vitamin K2 are interdependent. Vitamin C, selenium, and vitamin E are interdependent. Niacin and NAD are also dependent on adequate intracellular selenoproteins and vitamin C levels. Single nutrient studies and interventions will miss essential synergies and confounding variables regardless of the sample size. In critical illness, large doses of vitamin C can be helpful in resuscitation efforts. In the setting of septic vascular collapse, intravenous co-administration of vitamin C and cortisol helps the body repair damaged blood vessels quickly. Ongoing large doses of vitamin C are needed to fuel white blood cells, regain antioxidant capacity throughout the body, and counteract its rapid consumption. Frequent dosing to maintain a steady state is better, because ascorbic acid has a short half-life. Early intervention is better, because activated white blood cells are dependent on a high level of ascorbic acid. Taking gram quantities with each meal, and increasing intake to bowel tolerance during illness, is helpful. When ill, it is necessary to take ascorbic acid throughout the day, much more than can be absorbed in one sitting. New research techniques, and new viruses improve our understanding of biochemistry and biology, and reinforce a longstanding concern - inadequate nutrition remains global and public health enemy #1. Recommended adult doses to reduce risk of serious infection: [24-30]
(Michael E. Passwater, son of author and columnist Dr. Richard Passwater, is certified by the American Society for Clinical Pathology as a medical technologist, a specialist in immunohematology, and is a diplomate in laboratory management. He has worked in clinical laboratories for 29 years, and has previously written "Do the Math: "MATH+" Saves Lives" and "The VICTAS Trial: Designed to Fail," both published by the Orthomolecular Medicine News Service.) References1. Tomasa-Irriguible TM, Bielsa-Berrocal L (2021) "COVID-19: Up to| 82% critically ill patients had low Vitamin C values". Nutr J 20:66 https://pubmed.ncbi.nlm.nih.gov/34243781 2. Chiscano-Camón, L; Ruiz-Rodriguez, J.C.; Ruiz-Sanmartin, et al. (2020) Vitamin C levels in patients with SARS-CoV2-associated acute respiratory distress syndrome. Critical Care; 24:522. https://pubmed.ncbi.nlm.nih.gov/32847620 3. Carr AC, Rosengrave PC, Bayer S, et al. (2017) Hypovitaminosis C and vitamin C deficiency in critically ill patients despite recommended enteral and parenteral intakes. Critical Care, 21:300. https://pubmed.ncbi.nlm.nih.gov/29228951 4. Borrelli E, Roux-Lombard P, Grau GE, et al. (1996) Plasma concentrations of cytokines, their soluble receptors, and antioxidant vitamins can predict the development of multiple organ failure in patients at risk. Crit Care Med,24:392-397. https://pubmed.ncbi.nlm.nih.gov/8625625 5. Schleicher RL, Carroll MD, Ford ES, Lacher DA (2009) Serum vitamin C and the prevalence of vitamin C deficiency in the United States: 2003-2004 National Health and Nutrition Examination Survey (NHANES) . Am J Clin Nutr 2009;90:1252-1263. https://pubmed.ncbi.nlm.nih.gov/19675106 6. Carr AC, Maggini S. "Vitamin C and Immune Function". Nutrients 2017, 9, 1211; doi 10.3390/nu9111211 https://pubmed.ncbi.nlm.nih.gov/29099763 7. Buehler PW, Alayash AI (2005) Redox Biology of Blood Revisited: The Role of Red Blood Cells in Maintaining Circulatory Reductive Capacity. Antioxidants & Redox Signaling. 1755-1760. https://pubmed.ncbi.nlm.nih.gov/16356136 8. Wang Y, Zhao N, Xiong Y, et al. (2020) Downregulated Recycling Process but Not De Novo Synthesis of Glutathione Limits Antioxidant Capacity of Erythrocytes in Hypoxia. Oxidative Medicine and Cellular Longevity. 2020:7834252. https://pubmed.ncbi.nlm.nih.gov/32963701 9. Mayer JM, Marzinzig M, Kinzl L, et al. (2001) Reduced serum total reductive capacity in lethal severe trauma. J Trauma. 51(1):88-91. https://pubmed.ncbi.nlm.nih.gov/11468473 10. Dzik WH. (2011) The air we breathe: three vital respiratory gases and the red blood cell: oxygen, nitric oxide, and carbon dioxide. Transfusion 51: 676-685. https://pubmed.ncbi.nlm.nih.gov/21496039 11. Wang Y, Huang J, Sun Y, et al. (2021) SARS-CoV-2 suppresses mRNA expression of selenoproteins associated with ferroptosis, endoplasmic reticulum stress and DNA synthesis. Food and Chemical Toxicology 153 :112286. https://pubmed.ncbi.nlm.nih.gov/34023458 12. Thomas T, Stefanoni D, Dzieciatkowska M, et al. (2020) Evidence of Structural Protein Damage and Membrane Lipid Remodeling in Red Blood Cells from COVID-19 Patients. J Proteome Res. 19: 4455-4469. https://pubmed.ncbi.nlm.nih.gov/33103907 13. Aggarwal S, Acharjee A, Mukherjee A, et al. (2021) Role of Multiomics Data to Understand Host-Pathogen Interactions in COVID-19 Pathogenesis. J Proteome Res. 20:1107-1132. https://pubmed.ncbi.nlm.nih.gov/33426872 14. Khawaja UA, Shamsoddin E, Desideri LE, Tovani-Palone MR. (2021) Infection of red blood cells by SARS-CoV-2: new evidence. Einstein. (Sao Paulo) 2021 Apr 16;19:eCE6285. https://pubmed.ncbi.nlm.nih.gov/33886935 15. Shahbaz S, Xu L, Osman M, et al. (2021) Erythroid precursors and progenitors suppress adaptive immunity and get invaded by SARS-CoV-2. Stem Cell Reports, 16:1165-1181. https://pubmed.ncbi.nlm.nih.gov/33979601 16. Shi Y, Wang Y, Shao C, et al. (2020) COVID-19 infection: the perspectives on immune responses. Cell Death Differ. 27:1451-1454. https://pubmed.ncbi.nlm.nih.gov/32205856 17. Miller R, Wentzel AR, Richards GA. (2020) COVID-19: NAD+ deficiency may predispose the aged, obese and type2 diabetics to mortality through its effect on SIRT1 activity. Med Hypotheses, 144:110044. https://pubmed.ncbi.nlm.nih.gov/32758884 18. Taylor EW. (2010) The oxidative stress-induced niacin sink (OSINS) model for HIV pathogenesis PMID:19857540. Toxicology. 278:124-130. https://pubmed.ncbi.nlm.nih.gov/19857540 19. Polonikov A. (2020) Endogenous Deficiency of Glutathione as the Most Likely Cause of Serious Manifestations and Death in COVID-19 Patients. ACS Infect. Dis. 6:1558-1562. https://pubmed.ncbi.nlm.nih.gov/32463221 20. Hiffler L, Rakotoambinina B (2020) Selenium and RNA Virus Interactions: Potential Implications for SARS-CoV-2. Infection (COVID-19). Front. Nutr. 7:164. https://pubmed.ncbi.nlm.nih.gov/33015130 21. Vavougios GD, Ntoskas KT, Doskas TK. (2020) Impairment in selenocysteine synthesis as a candidate mechanism of inducible coagulopathy in COVID-19 patients. Med Hypotheses 147:110475. https://pubmed.ncbi.nlm.nih.gov/33421689 22. Moghaddam A, Heller RA, Sun Q, et al. (2020) Selenium Deficiency Is Associated with Mortality Risk from COVID-19. Nutrients, 12:2098. https://pubmed.ncbi.nlm.nih.gov/32708526 23. Horowitz RI, Freeman PR, Bruzzese J (2020) Efficacy of glutathione therapy in relieving dyspnea associated with COVID-19 pneumonia: A report of 2 cases. Respiratory Medicine Case Reports 30: 101063. https://pubmed.ncbi.nlm.nih.gov/32322478 24. Doctor Y, Saul AW, Smith RG (2021) Nutrition to Prevent and Treat COVID-19. Orthomolecular Medicine News Service. http://www.orthomolecular.org/resources/omns/v17n03.shtml (suggest adding niacin/niacinamide, K2 along with D3, and increased Se to 200 ug/day to the supplement recommendations listed) 25. Passwater M (2020) Do the Math: MATH+ Saves Lives. Orthomolecular Medicine News Service. http://www.orthomolecular.org/resources/omns/v16n55.shtml 26. Passwater M (2021) The VICTAS Trial: Designed to Fail.Orthomolecular Medicine News Service. http://www.orthomolecular.org/resources/omns/v17n08.shtml 27. Riordan Clinic (2021) High dose vitamin C (IVC) https://riordanclinic.org/what-we-do/high-dose-iv-vitamin-c 28. Holford P (2020) Vitamin C for the Prevention and Treatment of Coronavirus. Orthomolecular Medicine News Service. http://orthomolecular.org/resources/omns/v16n36.shtml 29. Gonzalez MJ (2020) Personalize Your COVID-19 Prevention: An Orthomolecular Protocol. Orthomolecular Medicine News Service. http://orthomolecular.org/resources/omns/v16n31.shtml 30. Rasmussen MPF (2020) Vitamin C Evidence for Treating Complications of COVID-19 and other Viral Infections. Orthomolecular Medicine News Service. http://orthomolecular.org/resources/omns/v16n25.shtml Nutritional Medicine is Orthomolecular MedicineOrthomolecular medicine uses safe, effective nutritional therapy to fight illness. For more information: http://www.orthomolecular.org Find a DoctorTo locate an orthomolecular physician near you: http://orthomolecular.org/resources/omns/v06n09.shtml The peer-reviewed Orthomolecular Medicine News Service is a non-profit and non-commercial informational resource. Editorial Review Board:
Albert G. B. Amoa, MB.Ch.B, Ph.D. (Ghana)
Comments and media contact: drsaul@doctoryourself.com OMNS welcomes but is unable to respond to individual reader emails. Reader comments become the property of OMNS and may or may not be used for publication. To Subscribe at no charge: http://www.orthomolecular.org/subscribe.html To Unsubscribe from this list: http://www.orthomolecular.org/unsubscribe.html |

This website is managed by Riordan Clinic
A Non-profit 501(c)(3) Medical, Research and Educational Organization
3100 North Hillside Avenue, Wichita, KS 67219 USA
Phone: 316-682-3100; Fax: 316-682-5054
© (Riordan Clinic) 2004 - 2024c
Information on Orthomolecular.org is provided for educational purposes only. It is not intended as medical advice.
Consult your orthomolecular health care professional for individual guidance on specific health problems.